lunes, 11 de junio de 2012
miércoles, 6 de junio de 2012
TAREA DE LA UNIDAD 8
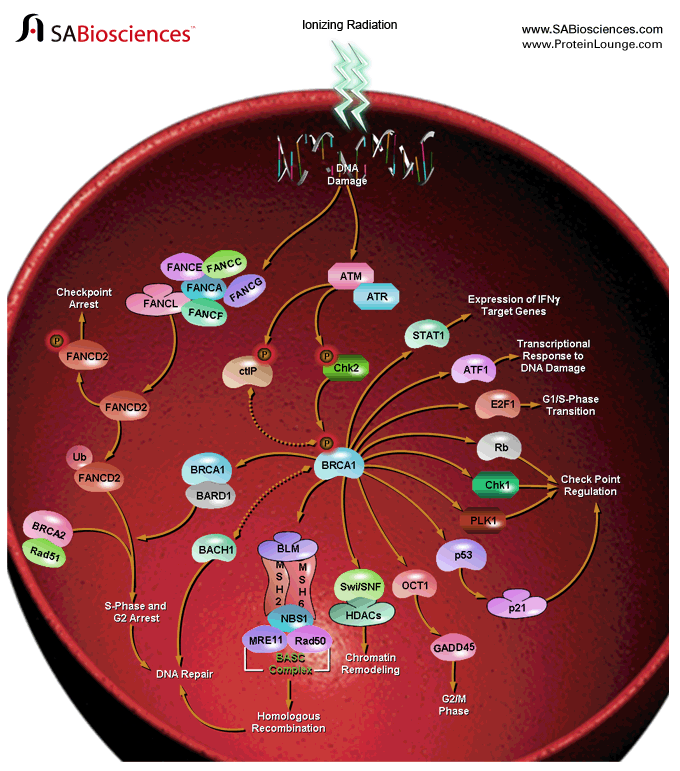
es un gen humano del tipo de los gen supresor de tumores, que regulan el ciclo celular y evitan la proliferación incontrolada. La proteína BRCA1, producto de este gen, forma parte del sistema de detección y reparación de los daños del ADN. Las variaciones de este gen están implicadas en algunos tipos de cáncer, especialmente el cáncer de mama. El gen BRCA1 está situado en el brazo largo (q) del cromosoma 17, en la posición 21, desde el par de bases 38.449.843 hasta el par de bases 38.530.933
Algunas variaciones del gen BRCA1 conducen a un mayor riesgo para generar cáncer de mama. Los investigadores han identificado más de 600 mutaciones en el gen BRCA1, muchos de los cuales están asociados con un mayor riesgo de cáncer.
Estas mutaciones pueden ser cambios en una o un pequeño número de pares de bases de ADN. Esas mutaciones pueden ser identificadas mediante la técnica de PCR y secuenciación del ADN.
En algunos casos, grandes segmentos de ADN se han reorganizado. Los grandes sectores, también llamados grandes reordenamientos, puede ser una deleción o una duplicación de uno o varios exones del gen. Los métodos clásicos para la detección de mutaciones (secuenciación) son incapaces de revelar las mutaciones. Se proponen otros métodos: Q-PCR, Multiplex Ligation-dependent Probe Amplification (MLPA), y Quantitative Multiplex PCR of Shorts Fluorescents Fragments (QMPSF). Los nuevos métodos se han propuesto recientemente: el análisis de heterodúplex (HDA) en electroforesis capilar o también hibridación genómica comparada (CGH array).
La participación de BRCA1 en el desarrollo del cáncer de mama se ha propuesto en varios estudios en los que está involucrada la hipermetilación de su promotor. Algunos resultados sugieren que la hipermetilación podría ser considerada como un mecanismo para inactivar la expresión de BRCA1, que ha sido reportada en algunos casos de cáncer.Un gen BRCA1 mutado normalmente produce una proteína que no funciona correctamente porque es anormalmente corta. Los investigadores creen que las proteínas defectuosas BRCA1 no está en condiciones de ayudar a corregir las mutaciones que se producen en otros genes. Estos defectos se acumulan y pueden permitir a las células crecer y dividirse de forma descontrolada, formando un tumor. Además de cáncer de mama, las mutaciones en el gen BRCA1 también aumentar el riesgo de cáncer de ovario, trompas de Falopio y de la próstata. Además, las lesiones precancerosas (displasia) dentro de la trompa de Falopio se han relacionado con mutaciones genéticas BRCA1.
tarea unidad IX
LAS BACTERIAS DE DIFERENTES ESPECIES PUEDEN COMPARTIR PLASMIDOS DE MANERA NATURAL?
SI , SI PUEDEN COMPARTIR PLASMIDOS LAS BACTERIAS DE DIFERENTES ESPECIES YA QUE TODAS LAS BACTERIAS TIENENCARACTERISTICAS SEMEJANTES , Y ALGUNOS PLASMIDOS NO RECONOCERAN LAS DIFERENCIAS Y ENTRARAN EN LAS BACTERIAS PROVOCANDO SU TRANSFORMACION PERO TAMBIEN EXISTEN PLASMIDOS ESPECIFICOS QUE SOLO PUEDEN ENTRAR EN LA PARED DE ALGUNAS BACTERIAS POR CARACTERISTICAS ESPECIFICAS DE ESAESPECIE DE BACTERIA.
lunes, 4 de junio de 2012
QUIMICOS
METODO DEL FOSFATO CALCICO
Uno de los métodos más baratos (y fiables) es la transfección mediante fosfato de calcio, originalmente descubierta por F. L. Graham y A. J. van der Eb en 1973. Una solución salina tamponada con HEPES y que contiene iones fosfato se combina con una solución de cloruro de calcio que contiene el DNA a transfectar. Cuando ambas se combinan, se forma un precipitado fino formado por el calcio cargado positivamente y el fosfato cargado negativamente, que el DNA en su superficie. Esta suspensión se añade a las células que se quieren transfectar (normalmente un cultivo celular en monocapa). Mediante un proceso no comprendido completamente , las células toman parte del precipitado, y junto con él, el DNA.
METODO DE LIPOSOMAS
Otros métodos usan compuestos orgánicos altamente ramificados, los llamados dendrímeros, para unir el DNA e introducirlo en la célula. Un método muy eficiente es la inclusión del DNA en liposomas, pequeños cuerpos formados de una membrana en cierto modo similar a la membrana plasmática de la célula y que puede fusionarse con la misma, liberando el DNA al interior celular. Con células eucariotas, la transfección basada en la interacción lípido-catión es la más comúnmente utilizada, ya que son más sensibles a este método.
METODO DEL DEAE DEXTRANO
Otro método es el uso de polímeros catiónicos (o policationes) como DEAE-dextrano o polietilenimina. El DNA, cargado negativamente, se une al policatión y el complejo es endocitado por la célula.
METODO DEL DNA DESNUDOEl DNA desnudo (técnica en fase altamente experiemtal) es incapaz de entrar en una célula y aún consiguiendo entrar en ellas es rápidamente degradado
METODO DE PEPTIDOS FUSIOGENICOS
Los péptidos fusiogénicos surgen en una linea de trabajo que nos permita paliar ... Estos péptidos fusiogénicosa se utilizan para compactar DNA
MECANISMOS DE TRANSFERENCIA ARTIFICIAL
FISICOS
microinyección es un proceso que consiste en utilizar microagujas para insertar sustancias a un nivel microscópico o en el límite de lo macroscópico dentro de una célula viva. Es un simple proceso mecánico en el cual una aguja extremadamente fina penetra la membrana celular y a veces la membrana nuclear para lanzar su contenido. La microinyección es normalmente realizada bajo un microscopio óptico llamado micromanipulador. El proceso es frecuentemente usado como un vector en ingeniería genética y transgenética para insertar material genético en una célula. El proceso de clonación también involucra microinyecciones.
Las microagujas miden alrededor de 10 micrómetros. Pueden contener cerca de 15 microlitros de ADN. Es similar a la sección transversal de un cabello humano. Es un método muy preciso de transferencia génica. Requiere personal capacitado.
electroporación o electropermeabilización es un significativo aumento de la conductividad eléctrica y la permeabilidad de la membrana plasmática celular causado por un campo eléctrico aplicado externamente. Es habitual en biología molecular como forma de introducción de diferentes sustancias en células, como por ejemplo sondas moleculares, un fármaco que puede cambiar las funciones celulares o un fragmento de DNA codificante, como puede ser un plásmido.
Cuando el voltaje que atraviesa una membrana plasmática excede su rigidez dieléctrica se forman poros. Si la fuerza del campo eléctrico aplicado o la duración de la exposición al mismo se eligen apropiadamente, los poros formados por el pulso eléctrico se sellan tras un corto período, durante el cual los compuestos extracelulares tienen la oportunidad de entrar a la célula. Sin embargo, una exposición excesiva de células vivas a campos eléctricos puede causar apoptosis o necrosis, procesos que provocan la muerte celular.
En biología molecular, el proceso de electroporación es usado habitualmente para la transformación de bacterias, levaduras y protoplastos vegetales. Además de membranas lipídicas, las bacterias también tienen una pared celular compuestas de peptidoglicano y sus derivados. Sin embargo, las paredes son porosas por naturaleza y sólo actúan como corazas que protegen a la célula de impactos ambientales severos. Si bacterias y plásmidos se mezclan los plásmidos pueden transferirse al interior de las células tras la electroporación. En este proceso suelen emplearse varios cientos de voltios, que atraviesan una distancia de varios milímetros. A continuación, las células han de ser manipuladas cuidadosamente hasta que tienen la oportunidad de dividirse, produciendo nuevas células que contendrán copias del plásmido. Este proceso es aproximadamente diez veces más efectivo que la transformación por métodos químicos.
Biobalistica:Una aproximación directa a la transfección es la biolística o gene gun (pistola génica), en la que el DNA se une a una nanopartícula compuesta de algún sólido inerte, generalmente oro, que es "disparada" directamente en el núcleo de la célula diana. El DNA puede también ser introducido en las células usando un virus como vector. En estos casos, la técnica es llamada transducción, y se dice que las células se transducen.
TRANSFECCION
La transfección consiste en la introducción de material genético externo en células eucariotas mediante plásmidos, vectores víricos (en este caso también se habla de transducción) u otras herramientas para la transferencia. El término transfección para métodos no virales se usa en referencia a células de mamífero, mientras que el término transformación se prefiere para describir las transferencias no virales de material genético en bacterias y células eucariotas no animales como hongos, algas o plantas.
La transfección de células animales generalmente se lleva a cabo abriendo poros o "agujeros" transitorios en la membrana plasmática de las células mediante electroporación, para permitir el paso del material genético (como construcciones de DNA superenrollado o siRNA) aunque pueden ser transfectadas incluso proteínas (como anticuerpos, por ejemplo). Además de la electroporación, se pueden utilizar otras técnicas para efectuar la transfección, como por ejemplo liposomas producidos mediante la mezcla de lípidos catiónicos con el material genético, que se fusionarán con la membrana plasmática celular y depositarán su carga adentro.
El significado original de "transfección" era "infección por transformación", es decir, introducción de DNA o RNA desde un virus procariótico ó bacteriófago en las células, resultando en una infección. Al tener el término transformación otro sentido en biología celular animal (un cambio genético que permite la propagación durante largos periodos de células en cultivo, o la adquisición de propiedades típicas de las células cancerígenas), el término transfección adquirió, para células animales, su actual signifiado de cambio en las propiedades celulares por la introducción de material genético.

TRANSDUCCION
La transducción es un proceso mediante el cual el ADN es transferido desde una bacteria a otra mediante la acción de un virus. También se utiliza para designar al proceso mediante el cual ADN exógeno es introducido en una célula mediante un vector viral. Esta es una herramienta que usualmente utilizan los biólogos moleculares para introducir en forma controlada un gen extraño en el genoma de una célula receptora.
Cuando los bacteriófagos (virus que infectan bacterias) infectan una célula bacteriana, su modo normal de reproducción consiste en capturar y utilizar la maquinaria de replicación, transcripción, y traducción de la célula de la bacteria receptora para producir gran cantidad de virones, o producir partículas virales, incluido el ADN o ARN viral y la cubierta de proteína.
CONJUGACION
CONJUGACION:
La conjugación bacteriana es el proceso de transferencia de información genética desde una célula bacteriana donadora a otra receptora. Este proceso fue descubierto por Joshua Lederberg y Edward Tatum en 1946. Este proceso es promovido por determinados tipos de plásmidos, que portan un conjunto de genes cuyos productos participan en el proceso, y que requiere contactos directos entre ambas células, con intervención de estructuras superficiales especializadas y de funciones específicas (pilus sexuales en los Gram negativos, y contacto íntimo en los Gram positivos).
Algunos de estos plásmidos se comportan como episomas, es decir, que pueden integrarse en el cromosoma; en este caso, si se produce la conjugación, se puede transferir el propio plásmido más un segmento adyacente del cromosoma, que a su vez podrá recombinarse con secuencias homólogas del cromosoma del receptor, dando lugar a un cromosoma híbrido

MECANISMOS DE TRANSFERENCIA NATURAL
TRANSFORMACION
TRANSFORMACION es la altertérminos para las alteraciones genéticas resultantes de introducir ADN por virus (transducción) o por contactos intercelulares entre bacterias (conjugación). A la transformación de células animales se le llama transfección.
El término transformación es también usado, de manera más general, para describir mecanismos de transferencia de ADN o ARN en biología molecular (es decir, teniendo en cuenta más que las consecuencias genéticas). Por ejemplo la producción de transgénicos como maíz transgénico requiere la inserción de nueva información genética en el genoma del maíz usando el mecanismo apropiado de transferencia de ADN; el proceso se le llama comúnmente transformación.
El ARN también puede ser transferido en las células usando métodos similares, pero esto no provoca normalmente cambios heredables y por lo tanto no es información real.

OBJETIVOS
CONOCER CUALES SON LOS MECANISMOS DE TRANSFERENCIA DEL MATERIAL GENETICO, NATURALES Y ARTIFICIALES.
CONOCER EN QUE CONSISTE CADA UNO DE LOS MECANISMOS( NATURALES Y ARTIFICIALES)
CONOCER EN QUE CONSISTE CADA UNO DE LOS MECANISMOS( NATURALES Y ARTIFICIALES)
PORTADA
INSTITUTO TECNOLOGICO DE CIUDAD ALTAMIRANO
BIOLOGIA MOLECULAR
UNIDAD
9
TRANSFERENCIA DEL MATERIAL GENETICO
ALUMNO:
MIGUEL ANGEL ARZATE TORRES
MAESTRO:
FRANCISCO JAVIER PUCHE ACOSTA
martes, 29 de mayo de 2012
CONCLUSIONES
EN LA UNIDAD NUMERO 8 OBSERVAMOS CUALES SON LOS PROCESOS NECESARIOS PARA QUE SE LLEVE A CABO LA SÍNTESIS DE PROTEÍNAS ASÍ COMO LOS NIVELES DE LA EXPRESION GENÉTICA EN ORGANISMOS PROCARIOTICOS Y EUCARIOTICOS ADEMAS DE COMO FUNCIONAN LOS MODELOS DEL OPERON LACTOSA Y TRIPTOFANO EN AUSENCIA Y EN PRESENCIA DE LA LACTOSA Y EL TRIPTOFANO. Y ADEMAS DE LAS DIFERENCIAS QUE OCURREN EN LOS DOS PROCESOS Y LAS RUTAS QUE SE TOMAN DE ACUERDO ALA PRESENCIA DE ALGUNOS FACTORES.
REGULACION DE LA EXPRESIÓN GENÉTICA EN ORGANISMOS EUCARIOTAS
Los organismos multicelulares complejos están compuestos de diferentes tejidos cuyas características individuales dependen de las proteínas específicas expresadas por sus tipos celulares. La diferenciación, el desarrollo y la funcionalidad de los tejidos específicos dependen del conjunto de proteínas selectivamente expresadas por cada célula. Estas proteínas expresadas en forma diferencial pueden funcionar como componentes estructurales de las células, enzimas reguladoras del metabolismo, factores de transcripción, receptores celulares, componentes intracelulares de señalización, etc.
La expresión incorrecta de tales proteínas, su expresión en lugares equivocados, a destiempo, o la producción en cantidades anormales de proteínas específicas o de proteínas de función anómala subyace a toda patología celular de base genética.
Por consiguiente el conocimiento de los mecanismos de regulación de la expresión proteica en eucariontes contribuirá al conocimiento de las bases moleculares de diversas patologías.
NIVELES DE REGULACIÓN DE LA EXPRESIÓN PROTEICA EN EUCARIONTES
En las células eucariotas, la capacidad de expresar proteínas biológicamente activas resulta de diferentes niveles regulatorios.
La compactación de la cromatina afecta la capacidad de unión de las enzimas y factores transcripcionales de genes específicos. La cromatina se puede dividir en dos clases según su patrón de tinción. La eucromatina se tiñe suavemente y se corresponde con regiones del genoma que están disponibles para la transcripción. Por otro lado, la heterocromatina, se tiñe intensamente y se corresponde a regiones del genoma que están densamente compactadas e inaccesibles para el aparato transcripcional.

Se pueden distinguir dos clases de heterocromatina: la constitutiva y la facultativa. La constitutiva hace referencia a cromosomas o parte de ellos que son heterocromáticos en todas las células de una misma especie, mientras que la facultativa implica zonas de cromosomas que se pueden descompactar tornándose en eucromatina en algunas células de un mismo organismo.
Como la heterocromatina no puede ser transcripta, la expresión génica en los eucariontes se puede reprimir por condensación de eucromatina en heterocromatina. Todavía no se conocen todos los factores que modulan la descompactación de la cromatina. Ciertamente hay proteínas que reconocen secuencias específicas del DNA y una vez unidas, transmiten la señal de descondensación de cerca de 10000 pares de bases correspondientes a un bucle de la cromatina.

Las acetilaciones y desacetilaciones de histonas son modificaciones covalentes frecuentes en estos fenómenos de descompactación cromatínica. Un ejemplo típico de este tipo de regulación ocurre en la acetilación de coactivadores involucrados en las transcripciones genéticas moduladas por las hormonas tiroideas. Las acetilaciones se producen en los residuos de lisina de los extremos aminoterminales de las histonas, reduciendo su carga positiva y por lo tanto su afinidad de unión al ADN cargado negativamente. La desacetilación de las histonas, mediada por desacetilasas provoca el efecto contrario (recompactación).
Secuencias características de organización del DNA como los palíndromes así como la disposición espacial del DNA Z han sido relacionados con señalizaciones para el sitio de inicio de la transcripción.
:
La metilación de los restos de citosina en el ADN, especialmente en los sitios promotores, dificultan la transcripción. Por ejemplo: los genes de globina están más metilados en células no productoras de hemoglobina que en los eritroblastos. Las metilaciones se producen en secuencias específicamente reconocidas ( 5’--- m CpG ---3’) que generalmente se agrupan en “islotes” ricos en GC, con frecuencia dentro o cerca de regiones reguladoras de la transcripción.
 La metilación puede inhibir la transcripción de los genes al interferir en la capacidad de los factores de transcripción para reconocer los sitios de unión al ADN o alterando las conformaciones del ADN dificultando la polimerización de la ARN polimerasa. Uno de los ejemplos más espectaculares de la metilación ocurre durante el fenómeno de impresión genómica. Así, el conjunto de cromosomas heredados del progenitor masculino no es funcionalmente equivalente al conjunto de cromosomas heredados de la madre.Existen por lo menos 100 genes sometidos a esta expresión diferencial. Las versiones activas e inactivas de los genes difieren en sus patrones de metilación. Las diferencias en los alelos se originan durante la gametogénesis.
La metilación puede inhibir la transcripción de los genes al interferir en la capacidad de los factores de transcripción para reconocer los sitios de unión al ADN o alterando las conformaciones del ADN dificultando la polimerización de la ARN polimerasa. Uno de los ejemplos más espectaculares de la metilación ocurre durante el fenómeno de impresión genómica. Así, el conjunto de cromosomas heredados del progenitor masculino no es funcionalmente equivalente al conjunto de cromosomas heredados de la madre.Existen por lo menos 100 genes sometidos a esta expresión diferencial. Las versiones activas e inactivas de los genes difieren en sus patrones de metilación. Las diferencias en los alelos se originan durante la gametogénesis.
med.unne.edu.ar/catedras/bioquimica/expresion.htm
lunes, 28 de mayo de 2012
8.4.2 OPERON TRIPTOFANO
El Operón Triptófano regula la transcripción de las enzimas que intervienen en una vía anabólica. Las cinco enzimas que regula este Operón pertenecen a la vía anabólica del aminoácido Triptófano.
Es más que innecesaria la síntesis de un aminoácido, por parte de la célula, cuando la misma contiene cantidades suficientes de él. Cuando la concentración de Triptófano es la adecuada el represor inactivo, codificado por el gen regulador, se une a otra molécula que le confiere la capacidad para acoplarse al operador. Al estar el operador ocupado, la ARN polimerasa no podrá formar el Complejo Promotor Abierto, por lo tanto, la síntesis de las enzimas intervinientes en la vía anabólica del Triptófano no son producidas. El co-represor, molécula que confiere la activación del represor, es nada más ni nada menos que el Triptófano.
 Es sumamente lógico que lo sea, ya que cuando hay una alta concentración de Triptófano, no es necesario gastar energía en sintetizarlo. Si la concentración de mencionado aminoácido es baja, el represor no podrá unirse al co-represor, por tal motivo el represor no adquirirá la capacidad de unirse al operador.
Es sumamente lógico que lo sea, ya que cuando hay una alta concentración de Triptófano, no es necesario gastar energía en sintetizarlo. Si la concentración de mencionado aminoácido es baja, el represor no podrá unirse al co-represor, por tal motivo el represor no adquirirá la capacidad de unirse al operador.
Al encontrarse el operador libre, la ARN polimerasa podrá iniciar la transcripción de los genes estructurales, obteniéndose las enzimas necesarias para poder sintetizar este aminoácido esencial.

8.2.1 OPERON LACTOSA
Un Operón es grupo de genes estructurales cuya expresión está regulada por los mismos elementos de control (promotor y operador) y genes reguladores.
Los principales elementos que constituyen un operón son los siguientes:Los genes estructurales: llevan información para polipéptidos. Se trata de los genes cuya expresión está regulada. Los operones bacterianos suelen contener varios genes estructurales, son poligénicos o policistrónicos. Hay algunos operones bacterianos que tienen un solo gene estructural. Los operones eucarióticos suelen contener un sólo gen estructural siendo monocistrónicos.
El promotor (P): se trata de un elemento de control que es una región del ADN con una secuencia que es reconocida por la ARN polimerasa para comenzar la transcripción. Se encuentra inmediatamente antes de los genes estructurales. Abreviadamente se le designa por la letra P.
El operador (O): se trata de otro elemento de control que es una región del ADN con una secuencia que es reconocida por la proteína reguladora. El operador se sitúa entre la región promotora y los genes estructurales. Abreviadamente se le designa por la letra O.
El gen regulador (i): secuencia de ADN que codifica para la proteína reguladora que reconoce la secuencia de la región del operador. El gen regulador está cerca de los genes estructurales del operón pero no está inmediatamente al lado. Abreviadamente se le denomina gen i.
Proteína reguladora: proteína codificada por el gen regulador. Está proteína se une a la región del operador.
Inductor: sustrato o compuesto cuya presencia induce la expresión de los genes.

En ausencia del inductor (la lactosa), la proteína represora producto del gen i se encuentra unida a la región operadora e impide la unión de la ARN-polimerasa a la región promotora y, como consecuencia, no se transcriben los genes estructurales

 En presencia del inductor (la lactosa), este se une a la proteína reguladora que cambia su conformación y se suelta de la región operadora dejando acceso libre a la ARN-polimerasa para que se una a la región promotora y se transcriban los genes estructurales. Por consiguiente, la presencia del inductor hace que se expresen los genes estructurales del operón, necesarios para metabolizar la lactosa
En presencia del inductor (la lactosa), este se une a la proteína reguladora que cambia su conformación y se suelta de la región operadora dejando acceso libre a la ARN-polimerasa para que se una a la región promotora y se transcriban los genes estructurales. Por consiguiente, la presencia del inductor hace que se expresen los genes estructurales del operón, necesarios para metabolizar la lactosa 8.2 REGULACION DE LA TRANSCRIPCION EN ORGANISMOS PROCARIOTICOS
http://www.ucm.es/info/genetica/grupod/Operon/Operon.htm

En las bacterias, a pesar de ser organismos unicelulares, también es necesario regular la expresión de los genes adaptándola a las necesidades ambientales. Es un principio de economía celular el que la expresión de los genes este regulada según las circunstancias celulares. Un buen ejemplo de esta situación en bacterias es la regulación de las enzimas implicadas en el metabolismo de los azúcares. Las bacterias pueden emplear para obtener energía distintas fuentes de carbono, como la glucosa, lactosa, galactosa, maltosa, ramnosa y xilosa. Existen enzimas capaces de introducir cada uno de estos azúcares en la bacteria y enzimas capaces de romperlos para obtener energía. Lógicamente, sería un despilfarro energético producir simultáneamente todos los enzimas necesarios para metabolizar los diferentes azúcares mencionados.

Por consiguiente, sería mucho más económico para la célula producir solamente las enzimas necesarias en cada momento, es decir, si en el medio en el que vive la bacteria la principal fuente de carbono es la lactosa, solamente se expresarían los genes necesarios para metabolizar la lactosa, mientras que los otros genes no se expresarían. Por tanto, es esencial que exista un mecanismo de regulación de la expresión génica, de manera que los genes se expresen cuando sea necesario.
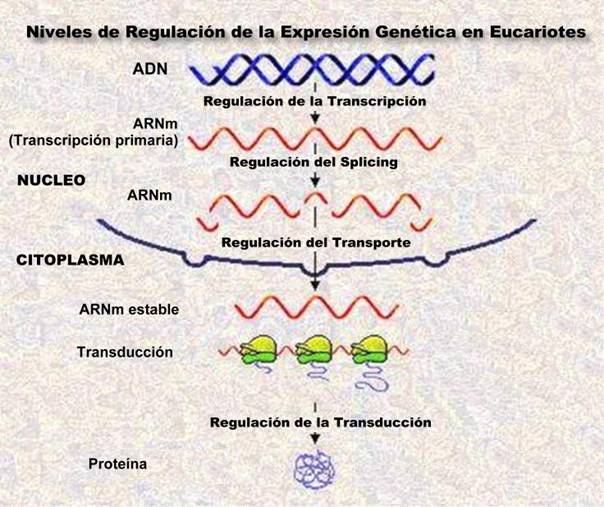
La regulación de la producción de proteínas (síntesis de proteínas) considerando el proceso en su conjunto, puede llevarse a cabo en tres niveles:
· Replicación
· Transcripción
· Traducción.
De los tres niveles de regulación, uno de los mejor conocidos actualmente es la regulación durante la transcripción.
8.1 NIVELES DE REGULACION DE LA EXPRESION GENETICA

El control de los genes regulables se realiza mediante proteínas que van a desarrollar un control activador o inhibidor sobre el mecanismo de la transcripción. Las células contienen un conjunto de proteínas que al unirse a secuencias específicas del ADN activan o desactivan los genes. Cada una de estas proteínas reguladoras de genes se encuentra en un número pequeño de copias y reconoce una secuencia de ocho a quince nucleótidos de la cadena del ADN. La unión puede facilitar (regulación positiva) o inhibir (regulación negativa) la transcripción de un gen adyacente.
En el caso de células procariotas, la mayoría de los ARNm son policistrónicos y pueden llevar transcritos de 2 a 6 genes. Los genes regulables que codifican proteínas de una ruta metabólica concreta no se encuentran dispersos en el genoma, sino que están normalmente adyacentes, agrupados en unidades de funcionamiento u operación denominadas operones, y su transcripción está bajo el control de proteínas activadoras y represoras. La región del ADN donde se unen estas proteínas recibe el nombre de operador y está muy próxima, si no solapada, con la región del promotor.
El ADN de Escherichia coli consta de un único cromosoma circular que contiene información para unas 4000 proteínas distintas; sin embargo, en un momento dado sólo se sintetizan algunas de ellas. Al ser un organismo procariota, regula la expresión de muchos de sus genes en función de los niveles intracelulares de metabolitos específicos, que varían según el medio ambiente que rodea a la célula. Los estudios genéticos sobre la utilización de lactosa como fuente alimenticia permitieron describir un modelo de regulación de expresión génica, el operón lactosa (lac), que es uno de los ejemplos mejor caracterizados de regulación a nivel de la transcripción.

Control positivo: Se dice que un sistema está bajo control positivo cuando el producto del gen regulador activa la expresión de los genes, actúa como un activador.
Control negativo: se dice que un sistema está bajo control negativo cuando el producto del gen regulador reprime o impide la expresión de los genes, actúa como un represor.
INTRODUCCION
REGULACION DE LA EXPRESION GENETICA

Los organismos multicelulares complejos están compuestos de diferentes tejidos cuyas características individuales dependen de las proteínas específicas expresadas por sus tipos celulares. La diferenciación, el desarrollo y la funcionalidad de los tejidos específicos dependen del conjunto de proteínas selectivamente expresadas por cada célula. Estas proteínas expresadas en forma diferencial pueden funcionar como componentes estructurales de las células, enzimas reguladoras del metabolismo, factores de transcripción, receptores celulares, componentes intracelulares de señalización, etc.
La expresión incorrecta de tales proteínas, su expresión en lugares equivocados, a destiempo, o la producción en cantidades anormales de proteínas específicas o de proteínas de función anómala subyace a toda patología celular de base genética.
Por consiguiente el conocimiento de los mecanismos de regulación de la expresión proteica en eucariontes contribuirá al conocimiento de las bases moleculares de diversas patologías.
NIVELES DE REGULACION DE LA EXPRESION PROTEICA EN EUCARIONTES
En las células eucariotas, la capacidad de expresar proteínas biológicamente activas resulta de diferentes niveles regulatorios.
La compactación de la cromatina afecta la capacidad de unión de las enzimas y factores transcripcionales de genes específicos. La cromatina se puede dividir en dos clases según su patrón de tinción. La eucromatina se tiñe suavemente y se corresponde con regiones del genoma que están disponibles para la transcripción. Por otro lado, la heterocromatina, se tiñe intensamente y se corresponde a regiones del genoma que están densamente compactadas e inaccesibles para el aparato transcripcional.
Se pueden distinguir dos clases de heterocromatina: la constitutiva y la facultativa. La constitutiva hace referencia a cromosomas o parte de ellos que son heterocromáticos en todas las células de una misma especie, mientras que la facultativa implica zonas de cromosomas que se pueden descompactar tornándose en eucromatina en algunas células de un mismo organismo.
Como la heterocromatina no puede ser transcripta, la expresión génica en los eucariontes se puede reprimir por condensación de eucromatina en heterocromatina. Todavía no se conocen todos los factores que modulan la descompactación de la cromatina. Ciertamente hay proteínas que reconocen secuencias específicas del DNA y una vez unidas, transmiten la señal de descondensación de cerca de 10000 pares de bases correspondientes a un bucle de la cromatina.
Las acetilaciones y desacetilaciones de histonas son modificaciones covalentes frecuentes en estos fenómenos de descompactación cromatínica. Un ejemplo típico de este tipo de regulación ocurre en la acetilación de coactivadores involucrados en las transcripciones genéticas moduladas por las hormonas tiroideas. Las acetilaciones se producen en los residuos de lisina de los extremos aminoterminales de las histonas, reduciendo su carga positiva y por lo tanto su afinidad de unión al ADN cargado negativamente. La desacetilación de las histonas, mediada por desacetilasas provoca el efecto contrario (recompactación).
Secuencias características de organización del DNA como los palíndromes así como la disposición espacial del DNA Z han sido relacionados con señalizaciones para el sitio de inicio de la transcripción.
Suscribirse a:
Comentarios (Atom)