Los organismos multicelulares complejos están compuestos de diferentes tejidos cuyas características individuales dependen de las proteínas específicas expresadas por sus tipos celulares. La diferenciación, el desarrollo y la funcionalidad de los tejidos específicos dependen del conjunto de proteínas selectivamente expresadas por cada célula. Estas proteínas expresadas en forma diferencial pueden funcionar como componentes estructurales de las células, enzimas reguladoras del metabolismo, factores de transcripción, receptores celulares, componentes intracelulares de señalización, etc.
La expresión incorrecta de tales proteínas, su expresión en lugares equivocados, a destiempo, o la producción en cantidades anormales de proteínas específicas o de proteínas de función anómala subyace a toda patología celular de base genética.
Por consiguiente el conocimiento de los mecanismos de regulación de la expresión proteica en eucariontes contribuirá al conocimiento de las bases moleculares de diversas patologías.
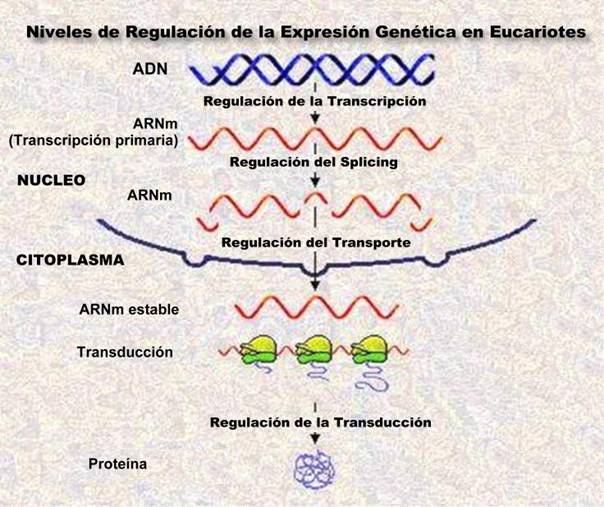
NIVELES DE REGULACION DE LA EXPRESION PROTEICA EN EUCARIONTES
En las células eucariotas, la capacidad de expresar proteínas biológicamente activas resulta de diferentes niveles regulatorios.
La compactación de la cromatina afecta la capacidad de unión de las enzimas y factores transcripcionales de genes específicos. La cromatina se puede dividir en dos clases según su patrón de tinción. La eucromatina se tiñe suavemente y se corresponde con regiones del genoma que están disponibles para la transcripción. Por otro lado, la heterocromatina, se tiñe intensamente y se corresponde a regiones del genoma que están densamente compactadas e inaccesibles para el aparato transcripcional.
Se pueden distinguir dos clases de heterocromatina: la constitutiva y la facultativa. La constitutiva hace referencia a cromosomas o parte de ellos que son heterocromáticos en todas las células de una misma especie, mientras que la facultativa implica zonas de cromosomas que se pueden descompactar tornándose en eucromatina en algunas células de un mismo organismo.
Como la heterocromatina no puede ser transcripta, la expresión génica en los eucariontes se puede reprimir por condensación de eucromatina en heterocromatina. Todavía no se conocen todos los factores que modulan la descompactación de la cromatina. Ciertamente hay proteínas que reconocen secuencias específicas del DNA y una vez unidas, transmiten la señal de descondensación de cerca de 10000 pares de bases correspondientes a un bucle de la cromatina.
Las acetilaciones y desacetilaciones de histonas son modificaciones covalentes frecuentes en estos fenómenos de descompactación cromatínica. Un ejemplo típico de este tipo de regulación ocurre en la acetilación de coactivadores involucrados en las transcripciones genéticas moduladas por las hormonas tiroideas. Las acetilaciones se producen en los residuos de lisina de los extremos aminoterminales de las histonas, reduciendo su carga positiva y por lo tanto su afinidad de unión al ADN cargado negativamente. La desacetilación de las histonas, mediada por desacetilasas provoca el efecto contrario (recompactación).
Secuencias características de organización del DNA como los palíndromes así como la disposición espacial del DNA Z han sido relacionados con señalizaciones para el sitio de inicio de la transcripción.
No hay comentarios:
Publicar un comentario