PLASMIDOS
Las moléculas de ADN plasmídico, adoptan una conformación tipo doble hélice al igual que el ADN de los cromosomas, aunque, por definición, se encuentran fuera de los mismos. Se han encontrado plásmidos en casi todas las bacterias. A diferencia del ADN cromosomal, los plásmidos no tienen proteínas asociadas.
En general, no contienen información esencial, sino que confieren ventajas al hospedador en condiciones de crecimiento determinadas. El ejemplo más común es el de los plásmidos que contienen genes de resistencia a un determinado antibiótico, de manera que el plásmido únicamente supondrá una ventaja en presencia de ese antibiótico.
Hay algunos plásmidos integrativos, es decir, que tienen la capacidad de insertarse en el cromosoma bacteriano. Estos rompen momentáneamente el cromosoma y se sitúan en su interior, con lo cual, automáticamente la maquinaria celular también reproduce el plásmido. Cuando ese plásmido se ha insertado se les da el nombre de episoma.
Los plásmidos se utilizan en ingeniería genética por su capacidad de reproducirse de manera independiente del ADN cromosomal así como también porque es relativamente fácil manipularlos e insertar nuevas secuencias genéticas.

BACTERIOFAGOS
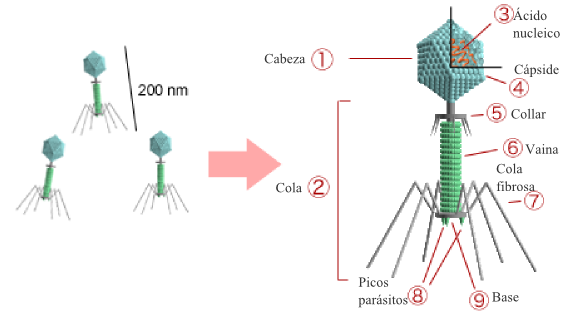
Los bacteriófagos (también llamados fagos -del griego φαγητόν (phagētón), «alimento, ingestión») son virus que infectan exclusivamente a bacterias.
Al igual que los virus que infectan células eucariotas, los fagos están constituidos por una cubierta proteica o cápside en cuyo interior está contenido su material genético, que puede ser ADN o ARN de simple o doble cadena, circular o lineal (en el 95% de los fagos conocidos es ADN de doble cadena), de 5.000 a 500.000 pares de bases. El tamaño de los fagos oscila entre 20 y 200 nm aproximadamente.
Los fagos son ubicuos y pueden ser encontrados en diversas poblaciones de bacterias, tanto en el suelo como en la flora intestinal de los animales. Uno de los ambientes más poblados por fagos y otros virus es el agua de mar, donde se estima que puede haber en torno a 109 partículas virales por mililitro, pudiendo estar infectadas por fagos el 70% de las bacterias marinas.
Los fagos pueden generar el ciclo lítico o el ciclo lisogénico, aunque muy pocos son capaces de llevar a cabo ambos. En el ciclo lítico, las células hospedadoras del fago son lisadas (destruidas) tras la replicación y encapsulación de las partículas virales, de forma que los nuevos virus quedan libres para llevar a cabo una nueva infección.
Por el contrario, en el ciclo lisogénico no se produce la lisis inmediata de la célula. El genoma del fago puede integrase en el ADN cromosómico de la bacteria hospedadora, replicándose a la vez que lo hace la bacteria o bien puede mantenerse estable en forma de plásmido, replicándose de forma independiente a la replicación bacteriana. En cualquier caso, el genoma del fago se transmitirá a toda la progenie de la bacteria originalmente infectada. El fago queda así en estado de latencia hasta que las condiciones del medio se vean deterioradas: disminución de nutrientes, aumento de agentes mutagénicos, etc. En este momento, los fagos endógenos o profagos se activan y dan lugar al ciclo lítico que termina con la lisis celular.

TRANSPOSONES
El transposón modifica el ADN de sus inmediaciones, ya sea arrastrando un gen codificador de un cromosoma a otro, rompiéndolo por la mitad o haciendo que desaparezca del todo. En algunas especies, la mayor parte del ADN basura (hasta un 50% del total del genoma) corresponde a transposones.
A diferencia de los provirus, los transposones se integran en el ADN celular en lugares bien determinados. Su existencia fue propuesta por Barbara McClintock en el maíz, sin embargo, su existencia no se demostró hasta mucho más tarde en bacterias. Por ello fue laureada con el Premio Nobel en 1983.
Clasificación
Existe una amplia diversidad de elementos genéticos móviles y pueden ser clasificados en base a su contenido y su estrategia y mecanismo de transposición.
Según contenido
- Transposón simple, secuencia de inserción o elemento de inserción (IS): contienen una secuencia central con información para la transposasa, una enzima necesaria para la transposición, y en los extremos una secuencia repetida en orden inverso. Esta secuencia repetida en orden inverso no es necesariamente idéntica, aunque muy parecida. Cuando un transposón simple se integra en un determinado punto del ADN aparece una repetición directa de la secuencia diana (5-12 pb).
- Transposón compuesto (Tn): contienen un elemento de inserción (IS) en cada extremo en orden directo o inverso y una región central con la transposasa que además suele contener información de otro tipo. Por ejemplo, los factores de transferencia de resistencia (RTF), poseen información en la zona central para resistencia a antibióticos como el cloranfenicol, la kanamicina, la tetraciclina, dándole una ventaja selectiva a las bacterias que lo posean.
Según estrategia de transposición
- Clase I o retrotransposones: se mueven en el genoma siendo transcritos a ARN y después en ADN por retrotranscriptasa. A su vez, se clasifican en los de origen retroviral (retrotransposones con LTR) y de origen no retroviral (retrotransposones sin LTR).
- Clase II o DNA transposones: se mueven directamente de una posición a otra en el genoma usando una transcriptasa para copiar y pegarse en otro locus del mismo.
- Clase III o MITE, por sus siglas en inglés "Miniature Inverted-repeats Transposable Elements".[
Según mecanismo de transposición
Transposición conservativa: el transposón sale de la sede donadora que queda vacía y se incorpora en una nueva sede (sede receptora). No aumenta el número de copias del transposón en el interior de la célula.
Se expresa la transposasa, y realiza dos cortes de doble cadena a la misma altura en el genoma donante, dejando aislado el transposón. A continuación localiza una secuencia diana (pongamos, ATGCA) en el genoma aceptor, y realiza un corte cohesivo. Tras eso une los extremos a los del transposón aislado, y la ADN Polimerasa de la célula rellena las zonas de cadena sencilla dejadas en la secuencia señal tras el corte cohesivo. Debido a esto, la secuencia señal queda duplicada. Queda, sin embargo, un hueco en el genoma donante, que puede ser letal si no se repara. Realmente, en este caso se habla más de recombinación que de transposición.
- Transposición no conservativa: en este caso la transposasa realiza un corte cohesivo no solo en la secuencia diana, sino también en el genoma donante, dejando un corte a cada lado del transposón. A continuación integra todo el genoma donante con el aceptor, mediante un curioso mecanismo que forma un intermediario llamado “estructura entrecruzada”. Esta estructura es resuelta por un segundo enzima, la resolvasa, que según cómo lo resuelva dará lugar a una de las siguientes transposiciones:
- Transposición no replicativa: el genoma donante se libera, dejando el integrón en el genoma receptor. Al igual que en la transposición conservativa, queda un hueco en el genoma donante, que puede ser letal si no se repara.
- Transposición replicativa: se produce una replicación desde los extremos 3’ del genoma aceptor, lo que acaba por duplicar el transposón, y produciendo un genoma mixto llamado “cointegrado”. A continuación la resolvasa rompe el cointegrado mediante una recombinación recíproca, que une los extremos del ADN aceptor original (ahora con una de las copias del integrón) y libera el genoma donante de nuevo con su transposón.
No hay comentarios:
Publicar un comentario