EN LA UNIDAD NUMERO 8 OBSERVAMOS CUALES SON LOS PROCESOS NECESARIOS PARA QUE SE LLEVE A CABO LA SÍNTESIS DE PROTEÍNAS ASÍ COMO LOS NIVELES DE LA EXPRESION GENÉTICA EN ORGANISMOS PROCARIOTICOS Y EUCARIOTICOS ADEMAS DE COMO FUNCIONAN LOS MODELOS DEL OPERON LACTOSA Y TRIPTOFANO EN AUSENCIA Y EN PRESENCIA DE LA LACTOSA Y EL TRIPTOFANO. Y ADEMAS DE LAS DIFERENCIAS QUE OCURREN EN LOS DOS PROCESOS Y LAS RUTAS QUE SE TOMAN DE ACUERDO ALA PRESENCIA DE ALGUNOS FACTORES.
martes, 29 de mayo de 2012
REGULACION DE LA EXPRESIÓN GENÉTICA EN ORGANISMOS EUCARIOTAS
Los organismos multicelulares complejos están compuestos de diferentes tejidos cuyas características individuales dependen de las proteínas específicas expresadas por sus tipos celulares. La diferenciación, el desarrollo y la funcionalidad de los tejidos específicos dependen del conjunto de proteínas selectivamente expresadas por cada célula. Estas proteínas expresadas en forma diferencial pueden funcionar como componentes estructurales de las células, enzimas reguladoras del metabolismo, factores de transcripción, receptores celulares, componentes intracelulares de señalización, etc.
La expresión incorrecta de tales proteínas, su expresión en lugares equivocados, a destiempo, o la producción en cantidades anormales de proteínas específicas o de proteínas de función anómala subyace a toda patología celular de base genética.
Por consiguiente el conocimiento de los mecanismos de regulación de la expresión proteica en eucariontes contribuirá al conocimiento de las bases moleculares de diversas patologías.
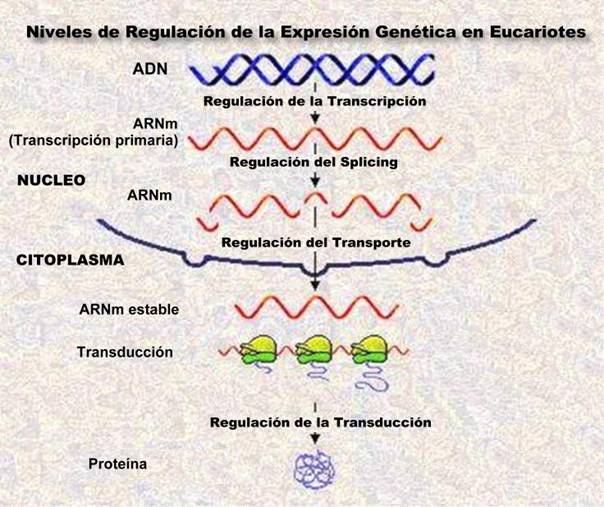
NIVELES DE REGULACIÓN DE LA EXPRESIÓN PROTEICA EN EUCARIONTES
En las células eucariotas, la capacidad de expresar proteínas biológicamente activas resulta de diferentes niveles regulatorios.
La compactación de la cromatina afecta la capacidad de unión de las enzimas y factores transcripcionales de genes específicos. La cromatina se puede dividir en dos clases según su patrón de tinción. La eucromatina se tiñe suavemente y se corresponde con regiones del genoma que están disponibles para la transcripción. Por otro lado, la heterocromatina, se tiñe intensamente y se corresponde a regiones del genoma que están densamente compactadas e inaccesibles para el aparato transcripcional.

Se pueden distinguir dos clases de heterocromatina: la constitutiva y la facultativa. La constitutiva hace referencia a cromosomas o parte de ellos que son heterocromáticos en todas las células de una misma especie, mientras que la facultativa implica zonas de cromosomas que se pueden descompactar tornándose en eucromatina en algunas células de un mismo organismo.
Como la heterocromatina no puede ser transcripta, la expresión génica en los eucariontes se puede reprimir por condensación de eucromatina en heterocromatina. Todavía no se conocen todos los factores que modulan la descompactación de la cromatina. Ciertamente hay proteínas que reconocen secuencias específicas del DNA y una vez unidas, transmiten la señal de descondensación de cerca de 10000 pares de bases correspondientes a un bucle de la cromatina.

Las acetilaciones y desacetilaciones de histonas son modificaciones covalentes frecuentes en estos fenómenos de descompactación cromatínica. Un ejemplo típico de este tipo de regulación ocurre en la acetilación de coactivadores involucrados en las transcripciones genéticas moduladas por las hormonas tiroideas. Las acetilaciones se producen en los residuos de lisina de los extremos aminoterminales de las histonas, reduciendo su carga positiva y por lo tanto su afinidad de unión al ADN cargado negativamente. La desacetilación de las histonas, mediada por desacetilasas provoca el efecto contrario (recompactación).
Secuencias características de organización del DNA como los palíndromes así como la disposición espacial del DNA Z han sido relacionados con señalizaciones para el sitio de inicio de la transcripción.
:
La metilación de los restos de citosina en el ADN, especialmente en los sitios promotores, dificultan la transcripción. Por ejemplo: los genes de globina están más metilados en células no productoras de hemoglobina que en los eritroblastos. Las metilaciones se producen en secuencias específicamente reconocidas ( 5’--- m CpG ---3’) que generalmente se agrupan en “islotes” ricos en GC, con frecuencia dentro o cerca de regiones reguladoras de la transcripción.
 La metilación puede inhibir la transcripción de los genes al interferir en la capacidad de los factores de transcripción para reconocer los sitios de unión al ADN o alterando las conformaciones del ADN dificultando la polimerización de la ARN polimerasa. Uno de los ejemplos más espectaculares de la metilación ocurre durante el fenómeno de impresión genómica. Así, el conjunto de cromosomas heredados del progenitor masculino no es funcionalmente equivalente al conjunto de cromosomas heredados de la madre.Existen por lo menos 100 genes sometidos a esta expresión diferencial. Las versiones activas e inactivas de los genes difieren en sus patrones de metilación. Las diferencias en los alelos se originan durante la gametogénesis.
La metilación puede inhibir la transcripción de los genes al interferir en la capacidad de los factores de transcripción para reconocer los sitios de unión al ADN o alterando las conformaciones del ADN dificultando la polimerización de la ARN polimerasa. Uno de los ejemplos más espectaculares de la metilación ocurre durante el fenómeno de impresión genómica. Así, el conjunto de cromosomas heredados del progenitor masculino no es funcionalmente equivalente al conjunto de cromosomas heredados de la madre.Existen por lo menos 100 genes sometidos a esta expresión diferencial. Las versiones activas e inactivas de los genes difieren en sus patrones de metilación. Las diferencias en los alelos se originan durante la gametogénesis.
med.unne.edu.ar/catedras/bioquimica/expresion.htm
lunes, 28 de mayo de 2012
8.4.2 OPERON TRIPTOFANO
El Operón Triptófano regula la transcripción de las enzimas que intervienen en una vía anabólica. Las cinco enzimas que regula este Operón pertenecen a la vía anabólica del aminoácido Triptófano.
Es más que innecesaria la síntesis de un aminoácido, por parte de la célula, cuando la misma contiene cantidades suficientes de él. Cuando la concentración de Triptófano es la adecuada el represor inactivo, codificado por el gen regulador, se une a otra molécula que le confiere la capacidad para acoplarse al operador. Al estar el operador ocupado, la ARN polimerasa no podrá formar el Complejo Promotor Abierto, por lo tanto, la síntesis de las enzimas intervinientes en la vía anabólica del Triptófano no son producidas. El co-represor, molécula que confiere la activación del represor, es nada más ni nada menos que el Triptófano.
 Es sumamente lógico que lo sea, ya que cuando hay una alta concentración de Triptófano, no es necesario gastar energía en sintetizarlo. Si la concentración de mencionado aminoácido es baja, el represor no podrá unirse al co-represor, por tal motivo el represor no adquirirá la capacidad de unirse al operador.
Es sumamente lógico que lo sea, ya que cuando hay una alta concentración de Triptófano, no es necesario gastar energía en sintetizarlo. Si la concentración de mencionado aminoácido es baja, el represor no podrá unirse al co-represor, por tal motivo el represor no adquirirá la capacidad de unirse al operador.
Al encontrarse el operador libre, la ARN polimerasa podrá iniciar la transcripción de los genes estructurales, obteniéndose las enzimas necesarias para poder sintetizar este aminoácido esencial.

8.2.1 OPERON LACTOSA
Un Operón es grupo de genes estructurales cuya expresión está regulada por los mismos elementos de control (promotor y operador) y genes reguladores.
Los principales elementos que constituyen un operón son los siguientes:Los genes estructurales: llevan información para polipéptidos. Se trata de los genes cuya expresión está regulada. Los operones bacterianos suelen contener varios genes estructurales, son poligénicos o policistrónicos. Hay algunos operones bacterianos que tienen un solo gene estructural. Los operones eucarióticos suelen contener un sólo gen estructural siendo monocistrónicos.
El promotor (P): se trata de un elemento de control que es una región del ADN con una secuencia que es reconocida por la ARN polimerasa para comenzar la transcripción. Se encuentra inmediatamente antes de los genes estructurales. Abreviadamente se le designa por la letra P.
El operador (O): se trata de otro elemento de control que es una región del ADN con una secuencia que es reconocida por la proteína reguladora. El operador se sitúa entre la región promotora y los genes estructurales. Abreviadamente se le designa por la letra O.
El gen regulador (i): secuencia de ADN que codifica para la proteína reguladora que reconoce la secuencia de la región del operador. El gen regulador está cerca de los genes estructurales del operón pero no está inmediatamente al lado. Abreviadamente se le denomina gen i.
Proteína reguladora: proteína codificada por el gen regulador. Está proteína se une a la región del operador.
Inductor: sustrato o compuesto cuya presencia induce la expresión de los genes.

En ausencia del inductor (la lactosa), la proteína represora producto del gen i se encuentra unida a la región operadora e impide la unión de la ARN-polimerasa a la región promotora y, como consecuencia, no se transcriben los genes estructurales

 En presencia del inductor (la lactosa), este se une a la proteína reguladora que cambia su conformación y se suelta de la región operadora dejando acceso libre a la ARN-polimerasa para que se una a la región promotora y se transcriban los genes estructurales. Por consiguiente, la presencia del inductor hace que se expresen los genes estructurales del operón, necesarios para metabolizar la lactosa
En presencia del inductor (la lactosa), este se une a la proteína reguladora que cambia su conformación y se suelta de la región operadora dejando acceso libre a la ARN-polimerasa para que se una a la región promotora y se transcriban los genes estructurales. Por consiguiente, la presencia del inductor hace que se expresen los genes estructurales del operón, necesarios para metabolizar la lactosa 8.2 REGULACION DE LA TRANSCRIPCION EN ORGANISMOS PROCARIOTICOS
http://www.ucm.es/info/genetica/grupod/Operon/Operon.htm

En las bacterias, a pesar de ser organismos unicelulares, también es necesario regular la expresión de los genes adaptándola a las necesidades ambientales. Es un principio de economía celular el que la expresión de los genes este regulada según las circunstancias celulares. Un buen ejemplo de esta situación en bacterias es la regulación de las enzimas implicadas en el metabolismo de los azúcares. Las bacterias pueden emplear para obtener energía distintas fuentes de carbono, como la glucosa, lactosa, galactosa, maltosa, ramnosa y xilosa. Existen enzimas capaces de introducir cada uno de estos azúcares en la bacteria y enzimas capaces de romperlos para obtener energía. Lógicamente, sería un despilfarro energético producir simultáneamente todos los enzimas necesarios para metabolizar los diferentes azúcares mencionados.

Por consiguiente, sería mucho más económico para la célula producir solamente las enzimas necesarias en cada momento, es decir, si en el medio en el que vive la bacteria la principal fuente de carbono es la lactosa, solamente se expresarían los genes necesarios para metabolizar la lactosa, mientras que los otros genes no se expresarían. Por tanto, es esencial que exista un mecanismo de regulación de la expresión génica, de manera que los genes se expresen cuando sea necesario.
La regulación de la producción de proteínas (síntesis de proteínas) considerando el proceso en su conjunto, puede llevarse a cabo en tres niveles:
· Replicación
· Transcripción
· Traducción.
De los tres niveles de regulación, uno de los mejor conocidos actualmente es la regulación durante la transcripción.
8.1 NIVELES DE REGULACION DE LA EXPRESION GENETICA

El control de los genes regulables se realiza mediante proteínas que van a desarrollar un control activador o inhibidor sobre el mecanismo de la transcripción. Las células contienen un conjunto de proteínas que al unirse a secuencias específicas del ADN activan o desactivan los genes. Cada una de estas proteínas reguladoras de genes se encuentra en un número pequeño de copias y reconoce una secuencia de ocho a quince nucleótidos de la cadena del ADN. La unión puede facilitar (regulación positiva) o inhibir (regulación negativa) la transcripción de un gen adyacente.
En el caso de células procariotas, la mayoría de los ARNm son policistrónicos y pueden llevar transcritos de 2 a 6 genes. Los genes regulables que codifican proteínas de una ruta metabólica concreta no se encuentran dispersos en el genoma, sino que están normalmente adyacentes, agrupados en unidades de funcionamiento u operación denominadas operones, y su transcripción está bajo el control de proteínas activadoras y represoras. La región del ADN donde se unen estas proteínas recibe el nombre de operador y está muy próxima, si no solapada, con la región del promotor.
El ADN de Escherichia coli consta de un único cromosoma circular que contiene información para unas 4000 proteínas distintas; sin embargo, en un momento dado sólo se sintetizan algunas de ellas. Al ser un organismo procariota, regula la expresión de muchos de sus genes en función de los niveles intracelulares de metabolitos específicos, que varían según el medio ambiente que rodea a la célula. Los estudios genéticos sobre la utilización de lactosa como fuente alimenticia permitieron describir un modelo de regulación de expresión génica, el operón lactosa (lac), que es uno de los ejemplos mejor caracterizados de regulación a nivel de la transcripción.

Control positivo: Se dice que un sistema está bajo control positivo cuando el producto del gen regulador activa la expresión de los genes, actúa como un activador.
Control negativo: se dice que un sistema está bajo control negativo cuando el producto del gen regulador reprime o impide la expresión de los genes, actúa como un represor.
INTRODUCCION
REGULACION DE LA EXPRESION GENETICA

Los organismos multicelulares complejos están compuestos de diferentes tejidos cuyas características individuales dependen de las proteínas específicas expresadas por sus tipos celulares. La diferenciación, el desarrollo y la funcionalidad de los tejidos específicos dependen del conjunto de proteínas selectivamente expresadas por cada célula. Estas proteínas expresadas en forma diferencial pueden funcionar como componentes estructurales de las células, enzimas reguladoras del metabolismo, factores de transcripción, receptores celulares, componentes intracelulares de señalización, etc.
La expresión incorrecta de tales proteínas, su expresión en lugares equivocados, a destiempo, o la producción en cantidades anormales de proteínas específicas o de proteínas de función anómala subyace a toda patología celular de base genética.
Por consiguiente el conocimiento de los mecanismos de regulación de la expresión proteica en eucariontes contribuirá al conocimiento de las bases moleculares de diversas patologías.
NIVELES DE REGULACION DE LA EXPRESION PROTEICA EN EUCARIONTES
En las células eucariotas, la capacidad de expresar proteínas biológicamente activas resulta de diferentes niveles regulatorios.
La compactación de la cromatina afecta la capacidad de unión de las enzimas y factores transcripcionales de genes específicos. La cromatina se puede dividir en dos clases según su patrón de tinción. La eucromatina se tiñe suavemente y se corresponde con regiones del genoma que están disponibles para la transcripción. Por otro lado, la heterocromatina, se tiñe intensamente y se corresponde a regiones del genoma que están densamente compactadas e inaccesibles para el aparato transcripcional.
Se pueden distinguir dos clases de heterocromatina: la constitutiva y la facultativa. La constitutiva hace referencia a cromosomas o parte de ellos que son heterocromáticos en todas las células de una misma especie, mientras que la facultativa implica zonas de cromosomas que se pueden descompactar tornándose en eucromatina en algunas células de un mismo organismo.
Como la heterocromatina no puede ser transcripta, la expresión génica en los eucariontes se puede reprimir por condensación de eucromatina en heterocromatina. Todavía no se conocen todos los factores que modulan la descompactación de la cromatina. Ciertamente hay proteínas que reconocen secuencias específicas del DNA y una vez unidas, transmiten la señal de descondensación de cerca de 10000 pares de bases correspondientes a un bucle de la cromatina.
Las acetilaciones y desacetilaciones de histonas son modificaciones covalentes frecuentes en estos fenómenos de descompactación cromatínica. Un ejemplo típico de este tipo de regulación ocurre en la acetilación de coactivadores involucrados en las transcripciones genéticas moduladas por las hormonas tiroideas. Las acetilaciones se producen en los residuos de lisina de los extremos aminoterminales de las histonas, reduciendo su carga positiva y por lo tanto su afinidad de unión al ADN cargado negativamente. La desacetilación de las histonas, mediada por desacetilasas provoca el efecto contrario (recompactación).
Secuencias características de organización del DNA como los palíndromes así como la disposición espacial del DNA Z han sido relacionados con señalizaciones para el sitio de inicio de la transcripción.
OBJETIVOS
Integrar los conocimientos anteriores con los mecanismos de regulación genética para entender a nivel molecular los procesos metabólicos.
PORTADA
miércoles, 23 de mayo de 2012
TAREA
EXPLIQUE: es posible que la maquinaria eucarionte
de traducción (ribosomas, factores, enzimas, RNAt) puedan traducir el RNAm de
una bacteria ?explique identificando los inconvenientes que encontrarías.
si es posible que
la maquinaria eucarionte pueda traducir el RNAm de una bacteria ya que el
proceso en eucariontes es un proceso más complejo, el ribosoma es más grande y
la hebra de ARNm debe de ser mas grande y como la de la bacteria seria mas pequeña
le facilitaría el proceso y seria mucho mas rápido siendo una hebra muy pequeña
para un ribosoma mas grande, además de que sobrarían enzimas y factores
de traducción y en este proceso como en cualquier otro si resulta un
producto o factor de mas si se podrá ,pero en cambio si falta un elemento
necesario para el proceso pues sería un proceso incompleto y no se podría
realizar.
viernes, 18 de mayo de 2012
7.4.1 MODIFICACIONES POSTRADUCCIONALES
La modificación postraduccional de una proteína es un cambio químico ocurrido en esta después de su síntesis proteica. Las modificaciones postraduccionales ocurren mediante cambios químicos de los aminoácidos que constituyen las proteínas y pueden ser de muchos tipos.
Modificaciones que añaden grupos funcionales
- Acilación
- Fosforilación
- Metilación
- Hidroxilación
- Glucosilación
- Sulfonilación
- Prenilación
- Glicación
- Nitrosilación
- Nitración
Modificaciones que enlazan proteínas
- Sumoilación
- Ubiquitinación: Consiste en la adición de la proteína ubiquitina a proteínas diana www.itescam.edu.mx/principal/sylabus/fpdb/recursos/r79527.PDF
miércoles, 16 de mayo de 2012
7.4 ETAPAS DE LA SINTESIS DE PROTEINAS EN ORGANISMOS EUCARIOTICOS
Eucariotas.
Es más complicada por:
- Muchos más factores de iniciación.
- No hay secuencias de Shine-Dalgarno, la colocación es por otros procedimientos, el tRNA no tiene una zona complementaria.
- Metionina no formilada.
La metionina unida al tRNA iniciador es lo único que se une a sólo una de las dos subunidades en el sitio P mal definido. Reacciona con EIF2. El primer aminoácido (metionina) se une con EIF2, cuando el ribosoma tiene el primer aminoácido se une al mensajero. El mRNA en eucariotas tiene gorra 5´, es el punto de enganche de la subunidad pequeña que ya tiene el primer aminoácido. Se desliza por el mensajero hasta encontrar el codon de iniciación y se para porque el tiene el aminoacil-tRNA que establece interacciones. Implica gasto de ATP porque hay estructuras que deshacer, hay factores proteicos que ayudan a deslizarse. Al entrar la subunidad grande se sueltan los factores proteicos.
Elongación.
Formación de los enlaces peptídicos desde el segundo aminoácido hasta el C-terminal. Ribosoma ya preparado. Entra el segundo aminoácido en el sitio A (la metionina está en el P) y se forma enlace peptídico. el ribosoma se desplaza, en el sitio P estarán la metionina y el segundo y en el sitio A el tercero. La entrada en el sitio A depende del codon.
En procariotas y eucariotas es casi igual. Hay tres factores de elongación:
| EF-Tu | EEF-1 α |
| EF-Ts | EEF-1βγ |
| EF-G | EEF-2 |
Un aminoácido para entrar en el ribosoma ha de estar unido a un factor proteico (EF-Tu o EEF-1 α) llamado aminoacil-tRNA:

El factor proteico nunca une ni tRNA iniciador ni tRNA sin aminoácido. El tRNA seleccionado según complementariedad dl codon.
Sitio P:

Los aminoácidos unidos por el carboxilo. Para formar peptídico el carboxilo 1° + amino 2°. Implica transferencia porque el CO que está activado se transfiere al α -amino participando una peptidiltransferasa.

En el sitio A queda un dipéptido unido por el tRNA del segundo. En el P queda tRNA vacío que se sale pasando por el sitio E. No necesita factor proteico extra. El RNA grande de la subunidad grande es la peptidiltransferasa. Tiene actividad catalítica. Para unir el siguiente aminoácido en el sitio A el tercer codon ha de estar desocupado, por lo que el complejo se ha de mover 3 nucleótidos (traslocación). Requiere factor proteico G activado con GTP. El factor proteico G ha de salir para que entre otro aminoácido porque sólo ayuda a moverse. Para salir hidroliza GTP. Se disocia porque solapa el sitio de Tu. El factor Ts sirve para reciclar Tu reactivándolo. Los factores se pueden usar varias veces si hay energía.
Terminación
Implica la disociación del complejo ribosoma + tRNA + proteína nueva.. Cuando en el sitio A aparezca un codon de parada tras el C-terminal o sin sentido, la peptidiltransferasa en lugar de transferir el peptídico a otro aminoácido lo transfiere al agua, gracias a los factores proteicos RF ó ERF
7.3 ETAPAS DE LA SINTESIS EN ORG.PROCARIOTICOS
La síntesis de proteínas ocurre en los ribosomas que consisten en dos subunidades, una grande y una pequeña, cada una formada por rRNA y proteínas específicas. Para la síntesis de proteínas, también se requiere de moléculas de tRNA, que están plegadas en una estructura secundaria con forma de hoja de trébol. Estas moléculas pequeñas pueden llevar un aminoácido en un extremo y tienen un triplete de bases, el anticodón, en un asa central, en el extremo opuesto de la molécula. La molécula de tRNA es el adaptador que aparea el aminoácido correcto con cada codón de mRNA durante la síntesis de proteínas. Hay al menos un tipo de molécula de tRNA para cada tipo de aminoácido presente en las células. Las enzimas conocidas como aminoacil-tRNA sintetasas catalizan la unión de cada aminoácido a su molécula de tRNA específica.
En E. coli y otros procariotas, aun cuando el extremo 3' de una cadena de mRNA está siendo transcripto, se están uniendo ribosomas cerca de su extremo 5'. En el lugar donde la cadena de mRNA está en contacto con un ribosoma, se unen tRNAs temporalmente a la cadena de mRNA. Esta unión ocurre por apareamiento de bases complementarias entre el codón de mRNA y el anticodón de tRNA. Cada molécula de tRNA lleva el aminoácido específico requerido por el codón de mRNA, al cual se une el tRNA. Así, siguiendo la secuencia dictada originalmente por el DNA, las unidades de aminoácidos son alineadas una tras otra y, a medida que se forman los enlaces peptídicos entre ellas, se unen en una cadena polipeptídica.
Esquema general del flujo de información en procariotas y eucariotas: a) En procariotas, el RNA se transcribe a partir de una molécula de DNA circular y, a medida que ocurre la transcripción, se produce la traducción en el mismo compartimiento. b) En eucariotas, la transcripción ocurre en el núcleo y el RNA, luego de sufrir un procesamiento, se dirige al citoplasma donde se produce la síntesis de proteínas. Como se vio en el capítulo 5, algunas proteínas son sintetizadas en los ribosomas libres y otras en los que están adheridos al retículo endoplásmico.
Tres etapas en la síntesis de proteínas en procariotas

La síntesis de proteínas ocurre en varias etapas: a) Iniciación. La subunidad ribosómica más pequeña se une al extremo 5' de una molécula de mRNA. La primera molécula de tRNA, que lleva el aminoácido modificado fMet, se acopla con el codón iniciador AUG de la molécula de mRNA. La subunidad ribosómica más grande se ubica en su lugar, el complejo tRNA-fMet ocupa el sitio P (peptídico). El sitio A (aminoacil) está vacante. El complejo de iniciación está completo ahora.

Un segundo tRNA, con su aminoácido unido, se coloca en el sitio A y su anticodón se acopla con el mRNA. Se forma un enlace peptídico entre los dos aminoácidos reunidos en el ribosoma. Al mismo tiempo, se rompe el enlace entre el primer aminoácido y su tRNA. El ribosoma se mueve a lo largo de la cadena de mRNA en una dirección 5' a 3', y el segundo tRNA, con el dipéptido unido, se mueve desde el sitio A al sitio P, a medida que el primer tRNA se desprende del ribosoma. Un tercer aminoacil-tRNA se coloca en el sitio A y se forma otro enlace peptídico. La cadena peptídica naciente siempre está unida al tRNA que se está moviendo del sitio A al sitio P y el tRNA entrante que lleva el siguiente aminoácido siempre ocupa el sitio A. Este paso se repite una y otra vez hasta que se completa el polipéptido.

Cuando el ribosoma alcanza un codón de terminación (en este ejemplo UGA), el polipéptido se escinde del último tRNA y el tRNA se desprende del sitio P. El sitio A es ocupado por un factor de liberación que produce la disociación de las dos subunidades del ribosoma.

A partir del DNA cromosómico se transcriben: diferentes moléculas de rRNA que, combinadas con proteínas específicas, forman los ribosomas; los diferentes tipos de moléculas de tRNA correspondientes a los distintos aminoácidos y los mRNA, que llevan la información para la secuencia de aminoácidos de las proteínas. Cuando un mRNA se une a la subunidad menor del ribosoma, comienza el proceso de síntesis de proteínas.
7.4 EUCARIOTICO
EUCARIOTAS

El ribosoma eucariota (32 nm) es más grande que el de procariotas, por lo que las subunidades de estos son de mayor tamaño y contienen más proteínas que sus análogos procariotas. En Eucariotas el 'coeficiente de sedimentación' S de los ribosomas completos es 80S, el cual está dividido en 60S para la grande y 40S para la subunidad pequeña. La subunidad mayor 60S contiene tres RNA ribosómicos (rRNA): 28S rRNA, 5S y 5.8S y unas 50 proteínas La subunidad mayor está conformada por tres ARN ribosomales (ARNr) de 25S, 5.8S y 5S, y 46 proteínas; mientras que la subunidad menor está formado sólo por un ARNr de 18S y 33 proteínas. Esto hace que el ribosoma eucariota sea un 40% grande que el ribosoma procariota.

La subunidad pequeña tiene solo un rRNA 18S y 33 proteínas. Los ribosomas eucariotas son sintetizados y ensamblados en el nucleolo . La formación del ribosoma es un proceso de autoensamblaje secuencial a partir de los rRNAs y de las proteínas ribosomales.
Los ribosomas se distribuyen en la célula eucariota en dos localizaciones: aquellos que están libres para deambular en el citosol (ribosomas citosólicos), y aquellos que están unidos a las membranas del retículo endoplasmático, dándole un aspecto granular visto al microscopio electrónico llamado por ello retículo endoplasmático rugoso (RER) . Aunque los dos clases de ribosomas tienen un papel similar en la síntesis de proteínas, se diferencian por la situación del producto final de su síntesis.

Los ribosomas en el citoplasma permiten que sus proteínas vagen libremente por el citosol mientras los ribosomas unidos al RER transfieren sus proteínas al lumen de este compartimento membranoso o la membrana del mismo donde pueden permanecer insertadas como proteínas de membrana dentro de la Vía Secretora . Además de los citados ribosomas, existen en la célula eucariota otros ribosomas: los mitocondriales y en las célula vegetales tienen además los ribosomas cloroplásticos.

7.2.3 PROCARIOTICO
PROCARIOTAS

En un ribosoma 70S, las dos subunidades se enfrentan de tal manera que se crea un espacio de morfologia complejo , y es uniformemente estrecha cuando se tiene una vision lateral. En la subunidad 30S se distinguen la cabeza y cuerpo. Del cuerpo salen dos lobulos hacia arriba, denominados “plataforma” y “espalda”. A la hendidura que se forma entre la plataforma y la cabeza se la conoce por ser el centro de decodificacion.
En bacterias, la subunidad 50S es aproximadamente dos veces la masa de la subunidad 30S, y en ambas, aproximadamente 2/3 de su peso se debe al ARNr mientras que el resto del peso es atribuible a la parte proteica del ribosoma.
La subunidad 30S tiene una forma mas o menos trapezoidal cuando se tiene una vista frontal desde la zona de interaccion de subunidades
La subunidad 50S tiene una forma aproximadamente hemisferica enfrentada a la subunidad 30S por su lado liso. Desde la region de interaccion entre subunidades se distinguen 3 protuberancias: el tallo L7/L12 que se extiende a lo largo de su base, la protuberancia central y el tallo L1. El tallo L7/L12, formado por las proteinas dimericas L7/L12, a veces puede estar ausente en mapas de microscopia electronica debido a su elevada flexibilidad. La protuberancia central esta formada, en su mayor parte, por el ARNr 5S. Al contrario que el tallo L7/L12, el tallo L1 esta siempre bien definido, formado por la proteina ribosomal L1 y parte del ARNr.

El espacio entre subunidades, por donde iran circulando los ARNt durante la traduccion. El ensamblaje se mantiene mediante diversos puentes que se crean entre subunidades. Existe un total de 12 puentes entre subunidades creados por mas de 30 interacciones individuales y la mayoria de ellos involucran contactos entre ARNr . Sin embargo, los puentes que se crean entre subunidades no confieren al ribosoma 70S una estructura totalmente rigida. La naturaleza dinamica del proceso de traduccion implica que el ribosoma sea una estructura flexible con componentes moviles que posibiliten su funcion.
www.argitalpenak.ehu.es/.../PATRICIA%20JULIAN%20TESIS.pdf
lunes, 14 de mayo de 2012
7.2.2 ESTRUCTURA RIBOSOMAL
RIBOSOMAS
.jpg)
Los ribosomas son los enzimas más complejos y de mayores dimensiones (aproximadamente 250Å de diámetro) que existen en la naturaleza. Están compuestos, incluso en los organismos más sencillos, de más de 50 proteínas distintas y ARNs que constituyen una masa por encima de los dos millones de daltons. Los ribosomas se componen de tres (en bacterias) o cuatro (en eucariotas) moléculas de ARNr y de tantas como 83 proteínas , organizadas en una subunidad mayor y en otra menor. Las subunidades ribosomales y las moléculas de ARNr se suelen designar en unidades Svedberg (S), una medida de la velocidad de sedimentación de partículas centrifugadas en condiciones estándares, de manera que en procariontes la subunidad menor se denomina 30S y la subunidad mayor 50S.

La subunidad 30S contiene una molécula única de ARNr, denominada ARNr 16S, mientras que la subunidad 50S contiene una molécula de 23S ARNr y otra 5S ARNr, La longitud de las moléculas de ARNr, la cantidad de proteínas en cada subunidad, y por consiguiente los tamaños de las subunidades son diferentes en células bacterianas y eucariotas. El ribosoma está constituido por el ensamblaje de las dos subunidades, que en eubacterias se denominan 70S y en eucariotas 80S. A pesar de que los ribosomas eucariotas contienen más componentes y son significativamente más grandes que los ribosomas procariotas, la apariencia general y el modus operandi es muy similar. La preservación de la función está unida con la conservación de su estructura general, y los ribosomas de organismos filogenéticamente tan lejanos como bacterias, arqueas y eucariotas muestran un grado remarcable de parecido.
Suscribirse a:
Entradas (Atom)